 Леонід ФАДЄЄВ
Леонід ФАДЄЄВ
Як відомо, насіння має складну форму і на плоских ситах, які використовуються для калібрування, розділяється за одним розміром - шириною. Але водночас кількість поживних речовин у зернівці в одній партії відкаліброваного насіння може сильно відрізнятися через різні значення його товщини, котра менша за ширину.
Винахід решіт нової геометрії дозволило відбирати насіння фактично за об’ємом. Різниця ширини і товщини у різних культурах різна і становить: у пшениці - 15 %, рисі - 20 %, гречці - 25 %, сої - 25 %, кукурудзі, просі, соняшнику - 30–35 %. На таких решетах легко відбирати за виповненістю будь-які культури, наприклад, соняшник, кукурудзу та ін. Крім того, на таких решетах легко і якісно відділяється сміття від насіння малих розмірів і товщини - зернівки повертаються, проходять крізь щілину решета, а сміття сходить із нього. Це можна побачити на прикладі насіння льону та іншого схожого за формою.
Легко відділяються половинки будь-яких бобових культур, наприклад, сої, залишати котрі з цілим насінням не можна, адже вони передусім вражаються грибами і стають провокаторами осередкового самозігрівання (їх дихання ушестеро інтенсивніше дихання цілого насіння сої).
 Значимість крупності насіння розглянемо на прикладі насіння пшениці. Після багатьох тисяч років землеробства ми отримали окультурені нашими пращурами рослини, потенціал яких зусиллями селекціонерів за останні 100 років вдалося суттєво підвищити. Але основний принцип відбору насіння для сівби залишився незмінним.
Значимість крупності насіння розглянемо на прикладі насіння пшениці. Після багатьох тисяч років землеробства ми отримали окультурені нашими пращурами рослини, потенціал яких зусиллями селекціонерів за останні 100 років вдалося суттєво підвищити. Але основний принцип відбору насіння для сівби залишився незмінним.
Проаналізуємо детальніше процес початку проростання рослин на прикладі пшениці. Як відомо, корінці пшениці діляться на два типи: зародкові (первинні) і придаткові (вторинні, стеблові). Зародкові корінці у зачатковому стані перебувають у складі зародку пшеничного зерна. Проростання зерна послідовно проходить через певні фази, першою з яких є фаза набухання, тобто поглинання зернівкою води. При набуханні зерна вода через оболонку потрапляє у колоїдні тканини зернівки і заповнює капіляри та міжклітковий простір.
Набухання - фізичний процес, інтенсивність якого залежить як від умов зовнішнього середовища, так і від хімічного складу зернівки. Необхідна кількість води, яку споживає зернівка пшениці для початку проростання, становить 43–44 % від її маси. Досліджуючи можливі шляхи отримання дружніх сходів, Є. Г. Кизилова (1961р.) виявила залежність інтенсивності набухання зерна пшениці від температури у насіннєвому ложі за оптимальної вологості грунту. Ця залежність експоненціальна - збільшення температури різко скорочується під час набухання. Крім того, поглинальна здатність складових зерна (крохмаль, білок, жир) відрізняється у рази. С. М. Богданов (1988р.), досліджуючи потребу насіння під час проростання у воді, переконливо показав цю різницю.
 Для проростання зерна це важливо, тому що ферменти для розщеплення вуглеводів, жирів та білків на прості форми цукрів, необхідні для живлення зародка, можуть діяти тільки у складі води. А оскільки ферменти активуються у зародку і алейроновому шарі під час їх руху до ендосперму, то висока здатність набухання білка, частка котрого в алейроновому шарі 30 %, а у зародку 26 %, сприяє активності процесу проростання на початковій фазі. Необерненість процесу проростання починається з моменту набухання головного зародкового корінця і, як наслідок, розриву оболонки та виходу його назовні. Наступна пара зародкових корінців проростає з певною затримкою після головного кореня. З появою волосків на зародкових корінцях вони починають забезпечувати паросток водою та поживними речовинами.
Для проростання зерна це важливо, тому що ферменти для розщеплення вуглеводів, жирів та білків на прості форми цукрів, необхідні для живлення зародка, можуть діяти тільки у складі води. А оскільки ферменти активуються у зародку і алейроновому шарі під час їх руху до ендосперму, то висока здатність набухання білка, частка котрого в алейроновому шарі 30 %, а у зародку 26 %, сприяє активності процесу проростання на початковій фазі. Необерненість процесу проростання починається з моменту набухання головного зародкового корінця і, як наслідок, розриву оболонки та виходу його назовні. Наступна пара зародкових корінців проростає з певною затримкою після головного кореня. З появою волосків на зародкових корінцях вони починають забезпечувати паросток водою та поживними речовинами.
Зародок дає початок життя не тільки корінцям, але й зародковому стеблу, першим прикореневим листочкам, адже вони, як і первинні корінці, перебувають у зародковому стані у складі зародка.
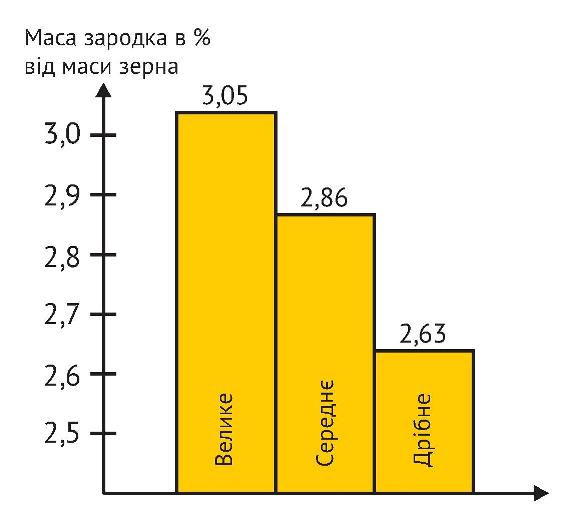
Таким чином, початок росту рослини відбувається тільки за рахунок витрачання поживних речовин, що перебувають в ендоспермі. Вони розкладаються ферментами до простих форм і у рідкій фазі через щиток потрапляють у зародок для розвитку первинної кореневої системи і зародкового стебла. Саме тому важливу роль грає кількість поживних речовин, тобто розмір та щільність ендосперму зернівки.
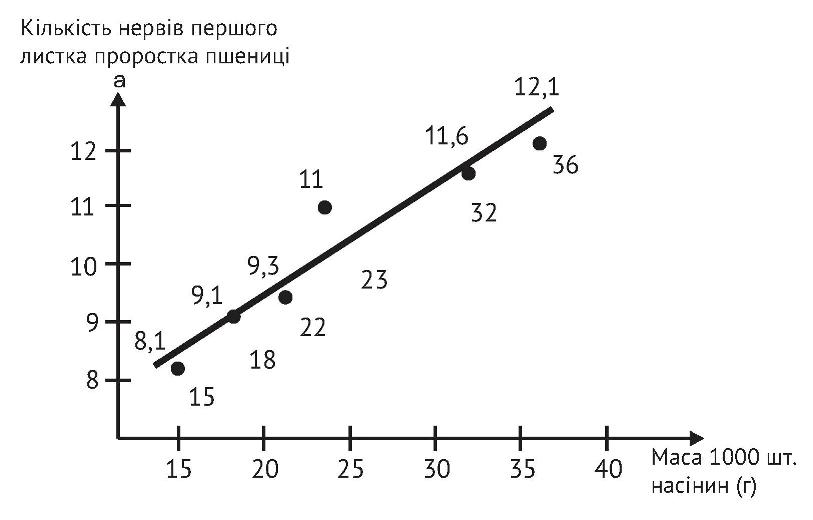
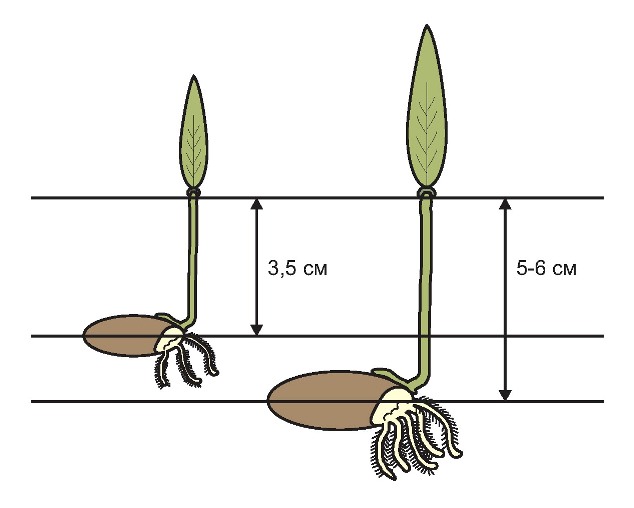 У крупного зерна не тільки крупніший зародок, що природно, але до того ж він крупніший навіть під час відносного порівняння. Так, І. Є. Мамбиш (1953 р.), досліджуючи вагові співвідношення складових частин пшениці та визначаючи масу зародку у дрібних та крупних зернах пшениці, як в абсолютній величині, так і у відносній, показав переваги крупного насіння. Саме тому крупне важке насіння пшениці дає вирівняні потужні сходи, оскільки первинні (зародкові) корінці і перший листок формуються практично тільки за рахунок поживних речовин сім’янки. Потужність зародкових корінців та площа першого листка напряму залежать від її крупності. А. І. Носатовський у книзі «Пшениця. Біологія» (1965р.) наводить дані, які переконливо підтверджують цю залежність.
У крупного зерна не тільки крупніший зародок, що природно, але до того ж він крупніший навіть під час відносного порівняння. Так, І. Є. Мамбиш (1953 р.), досліджуючи вагові співвідношення складових частин пшениці та визначаючи масу зародку у дрібних та крупних зернах пшениці, як в абсолютній величині, так і у відносній, показав переваги крупного насіння. Саме тому крупне важке насіння пшениці дає вирівняні потужні сходи, оскільки первинні (зародкові) корінці і перший листок формуються практично тільки за рахунок поживних речовин сім’янки. Потужність зародкових корінців та площа першого листка напряму залежать від її крупності. А. І. Носатовський у книзі «Пшениця. Біологія» (1965р.) наводить дані, які переконливо підтверджують цю залежність.
Наступні листки, включно до четвертого, формуються за рахунок двох джерел - із поживних речовин, які надходять через зародок від зернівки, і з зародкових корінців, які почали свою «роботу». Після того, як використано поживні речовини зерна, подальший розвиток рослини відбувається за рахунок зародкових корінців, адже розвиток придаткових корінців, за даними автора М. Г. Пруцкової (1976), у сприятливий рік починається приблизно через 18 днів після сходів, а у посушливий - через 28 днів.
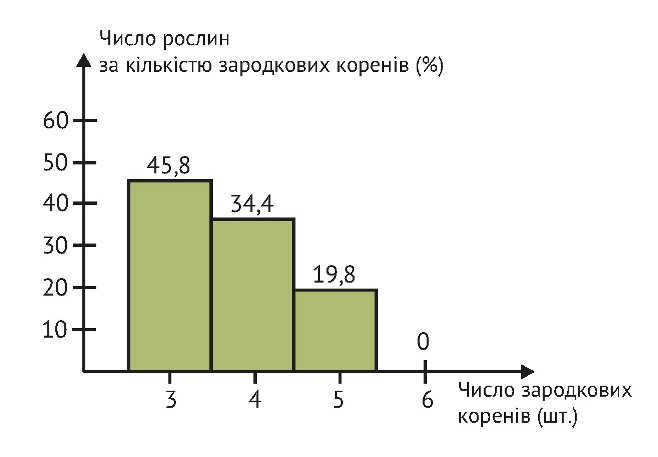
 Зародкові корінці швидко ростуть углиб і, досягнувши глибини 71–100 см, виявляються глибшими придаткових удвічі. Крім того, вони не відмирають за появи і розвитку вторинної кореневої системи і супроводжують всі основні фази розвитку рослин аж до молочно-воскової стиглості сформованого у колосі зерна. У зв’язку з цим знову повертаємося до значимості крупного насіння пшениці, тому що його виключно важлива властивість у тому, що кількість зародкових корінців у крупних насінин більше, ніж у дрібних. Д. Д. Ромащенков (1951р), досліджуючи залежність енергії проростання ярої пшениці від появи первинних корінців, наводить дані про те, що в рамках його досліджень понад 80 % крупних насінин було по 5 зародкових корінців, а у 80 % дрібних насінин - по 3–4 зародкових корінця.
Зародкові корінці швидко ростуть углиб і, досягнувши глибини 71–100 см, виявляються глибшими придаткових удвічі. Крім того, вони не відмирають за появи і розвитку вторинної кореневої системи і супроводжують всі основні фази розвитку рослин аж до молочно-воскової стиглості сформованого у колосі зерна. У зв’язку з цим знову повертаємося до значимості крупного насіння пшениці, тому що його виключно важлива властивість у тому, що кількість зародкових корінців у крупних насінин більше, ніж у дрібних. Д. Д. Ромащенков (1951р), досліджуючи залежність енергії проростання ярої пшениці від появи первинних корінців, наводить дані про те, що в рамках його досліджень понад 80 % крупних насінин було по 5 зародкових корінців, а у 80 % дрібних насінин - по 3–4 зародкових корінця.
Роль зародкових коренів у формуванні колоса озимої та ярової пшениці різна. Якщо в озимої пшениці вторинні корені з’являються восени і до колосіння досягають великої глибини, що і забезпечує урожай бокових пагонів, близький до урожаю головних пагонів, то у ярої пшениці картина інша. Вторинні корені формуються пізніше зародкових на 25–35 днів, залягають дрібніше, і основне навантаження на формування урожаю покладається на зародкові корені. Так, дослідження А. І. Носатовського показали, що частка урожайності, яка забезпечується зародковими коренями, становить не менше 70 % від урожаю, сформованого усією кореневою системою рослини. Якщо припустити, що наведені дані для порівняння продуктивності зародкових коренів належать до середньої їх кількості (3–4 шт.), то кількість зародкових коренів крупного насіння (5–6 шт.) можуть забезпечити ще більшу продуктивність за відсутності вторинних коренів.
 Особливо велика роль зародкових коренів для ярої пшениці (пшениці твердих сортів) у посушливі роки, коли у сухому шарі грунту вторинні корені не розвиваються, і весь урожай формується тільки за рахунок зародкових коренів. У цьому випадку крупне насіння, по суті, є засобом зниження втрат урожаю через посуху за рахунок великої кількості зародкових коренів, які уходять у грунт на глибину до 2000 см і більше в той час, коли стеблові корені у верхньому зневодненому шарі грунту припиняють свою життєдіяльність.
Особливо велика роль зародкових коренів для ярої пшениці (пшениці твердих сортів) у посушливі роки, коли у сухому шарі грунту вторинні корені не розвиваються, і весь урожай формується тільки за рахунок зародкових коренів. У цьому випадку крупне насіння, по суті, є засобом зниження втрат урожаю через посуху за рахунок великої кількості зародкових коренів, які уходять у грунт на глибину до 2000 см і більше в той час, коли стеблові корені у верхньому зневодненому шарі грунту припиняють свою життєдіяльність.
З цього також виходить, що крупна сім’янка пшениці з великим потенціалом і високою енергією проростання та великою кількістю висхідних поживних речовин у своєму складі може надійно проростати з більшої глибини закладення насіння при сівбі, що знижує ризики вимерзання для озимих культур і підвищує польову схожість за дефіциту вологи в період висіву, що особливо важливо для ярих твердих сортів пшениці, адже набухання склоподібних зерен відбувається повільно, і у цій фазі вони повинні перебувати у вологому грунті.
Зрозуміло, що крупність насіння - це половина справи, а ось друга половина - їх щільність. Про це піде мова наступного разу.
